This monograph will address several central questions. What are the uremic toxins, and from where do they originate? What role does the gut microbiome play In uremic toxicity and the progression of chronic kidney disease (CKD) and cardiovascular disease? How does diet affect the gut microbiome, and downstream, affect uremic toxicity?
The term uremia, literally meaning “urine in the blood,” was coined by the French physicians Pierre-Adolphe Piorry and Dominique L’Héritier in the 1840s to describe the constellation of signs and symptoms of advanced kidney failure. Urea is the most prevalent chemical found in urine. Interestingly, It has a place in history as the first organic compound synthesized from inorganic components, in 1828, by the German chemist Friedrich Wöhler. This achievement profoundly undermined the doctrine of “vitalism”, the prevailing belief that organic substances could only be produced by living organisms through a mysterious “vital force.”
To better understand the roles of urea and ammonia in nitrogen metabolism and balance, it is useful to follow the fate of nitrogen as it enters and leaves the body. Dietary protein Is digested and metabolized into amino acids and small peptides. Once in hepatocytes, some of these amino acids are deaminated so their carbon backbones can be used as energy and to form glucose. The remaining amino acids are used to form new proteins and other nitrogen-containing metabolites in the liver and in the rest of the body. To maintain nitrogen balance , the amount of nitrogen excreted must equal the amount absorbed in the GI tract. In other words, nitrogen-in equals nitrogen-out. As it turns out, 80% of excreted nitrogen is urea, which is formed from ammonia in liver hepatocytes, enters the systemic circulation, and is freely filtered into the urine by the kidneys.
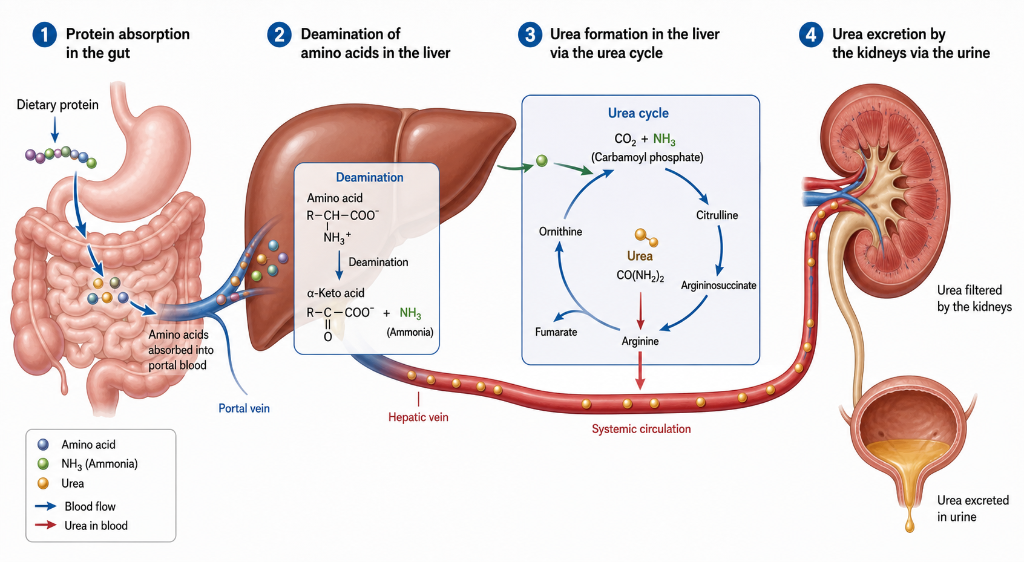
The urea cycle evolved over 200 million years and is conserved in all mammals, even though the conversion of ammonia to urea comes at an energy cost. Evolution took this path in part because of the dramatic difference in toxicity between ammonia and urea. The relatively non-toxic nature of urea is supported by a study where healthy volunteers were given hundreds of grams per hour of urea, raising their serum urea to levels comparable with severe uremia, with no untoward effects. (Eknoyan et al, NEJM, 1969) In another study, patients with end-stage renal disease on maintenance dialysis were dialyzed against a bath containing urea which maintained their serum urea at extremely high levels that would normally be consistent with uremic toxicity. Other small solutes were removed via dialysis as normal. These patients experienced no obvious signs of uremic toxicity or other negative effects. (Johnson et al, Mayo Clin Proc, 1972)
The fact that urea, the most prevalent substance in urine and the key to mammalian nitrogen balance, seems to be an underwhelming toxin leads to the question of identifying the true uremic toxins. Roughly 140 solutes have been identified that are retained in the uremic state. Keep in mind that the majority of these retained solutes are bystanders and not toxins. About one-third are small and water soluble (less than 500 Da), like urea, creatinine, and uric acid. These are freely filtered by the kidneys and are also efficiently cleared by conventional hemodialysis. Another one-third are small (less than 500 Da), largely protein-bound, and relatively water insoluble. The remaining one-third are middle-to-large in size (over 500 Da to over 50,000 Da). The vast majority are water soluble, and include β2-microglobulin, PTH, and various cytokines, like IL-1beta, IL-6, TNF-alpha. The most toxic compounds are found In the small and water insoluble group.
Before getting into specific toxins, two studies are notable because they establish the role of gut microbial flora in uremic toxicity. The first study compared plasma from dialysis patients who had undergone colectomy with control dialysis patients with intact colons. They identified the chemical signatures of over 30 compounds present in plasma from control patients but not colectomy patients. Five of these 30 compounds were known uremic toxins, and the other 25 unidentified compounds clearly accumulated and may well have also been toxins. (Aronov et al. JASN, 2011) The second study compared the survival of germ-free rats with control rats after undergoing bilateral nephrectomies. The germ-free rats lived almost twice as long as the control rats, strongly suggesting a role of the microbiome in accelerating the progression of uremia. (Einheber et al. J Exp Med, 1966)
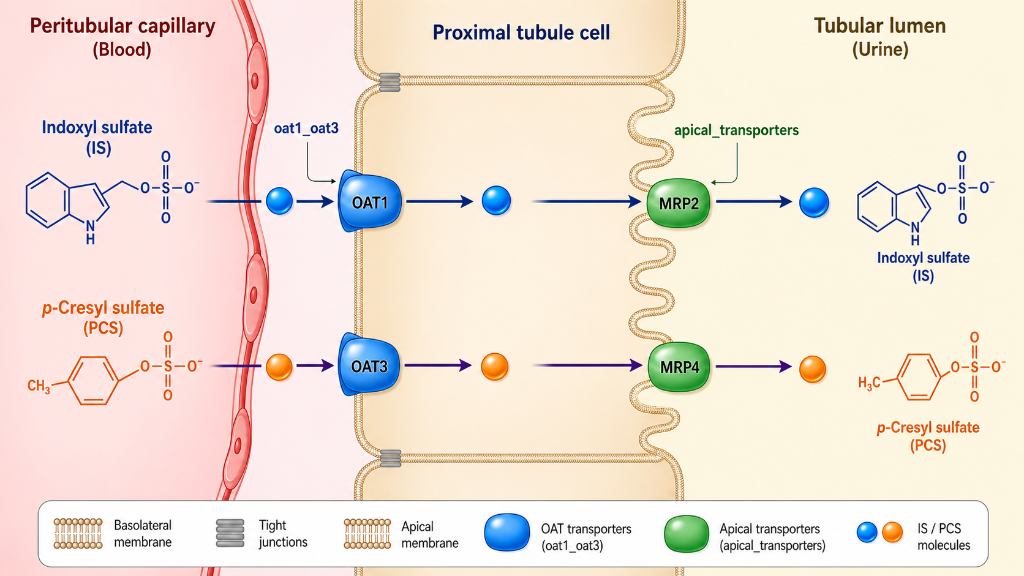
A description of three potent uremic toxins follows. All three are considered metaorganismal, meaning they cannot be formed In humans without some biochemical steps being performed by gut microbes. The first is indoxyl sulfate (IS). This is the prototypical gut-derived uremic toxin. Its formation begins when dietary tryptophan reaches the colon and Is cleaved by bacterial tryptophanase to produce free Indole. This indole is absorbed into the portal circulation and taken up by hepatocytes in the liver. Here, it is oxidized to indoxyl, then conjugated to form indoxyl sulfate, which is then released into the systemic circulation. Keeping in mind that it is highly protein bound, It is not freely filtered in the kidney and must be cleared through active secretion In the proximal tubules. Indoxyl sulfate causes widespread inflammation and scarring. It activates pro-inflammatory pathways in renal tubular cells, endothelial cells, and vascular smooth muscle cells. It promotes the generation of reactive oxygen species, causing further cell damage and inflammation. In addition, it promotes the formation of pro-inflammatory cytokines, such as ICAM-1 and IL-6.
Second is p-cresyl sulfate (PCS), which originates from gut bacterial metabolism of tyrosine and phenylalanine. Bacterial deaminases and decarboxylases convert these amino acids, through 4-hydroxyphenylacetic acid, to p-cresol, which enters the portal circulation. Once in the liver, p-cresol Is converted to PCS, which then enters the systemic circulation. PCS promotes adhesion of leukocytes to endothelial cells, promotes renal fibrosis, and increases Insulin resistance in skeletal muscle. Increased levels independently predict cardiovascular morbidity and mortality in CKD.
Diet has a large impact on IS and PCS production and levels. Plant based diets reduce IS and PCS production by 60%. In CKD patients, six weeks of fiber supplementation reduced free IS and PCS levels by roughly 25%. The protein-to-fiber ratio is the single best predictor of IS and PCS levels. The combined dietary pattern of high animal protein with low fiber, plus CKD, produces the highest toxin levels; while low animal protein with high fiber, plus normal kidneys, produces the lowest. Like IS, PCS is highly protein bound and is cleared by the kidneys via active secretion in the proximal tubules.
Third is trimethylamine N-oxide (TMAO). Like IS and PCS, TMAO is formed by metaorganismal pathways from several dietary precursors. The most prevalent precursor choline, found primarily in the form of phosphatidylcholine or lecithin, which is present in high concentrations in egg yolks and animal meat. The second most prevalent TMAO precursor is L-carnitine, which is found mainly in beef and pork.
Another source of TMAO is direct absorption of preformed TMAO found in fish. This is generally considered less impactful, as this TMAO is rapidly absorbed and cleared by the kidneys, as opposed to the slow, steady, constant production from dietary precursors metabolized by gut microbes. Getting back to the biochemical pathway of TMAO formation, gut bacteria metabolize choline, phosphatidylcholine, and L-carnitine to form TMA, or trimethylamine. TMA is absorbed through the colon and enters liver hepatocytes via the portal circulation, where it is metabolized to TMAO and released into the systemic circulation.
TMAO accelerates atherogenesis by altering lipid metabolism in macrophages and hepatocytes, impairing cholesterol efflux via macrophages in the periphery, and impairing bile acid synthesis in the liver. It also causes increased Intracellular calcium which induces platelet hyperreactivity, causing a hypercoagulable state. TMAO causes vascular endothelial dysfunction through a variety of mechanisms. These include upregulation of cytokines and adhesion molecules and increased production of reactive oxygen species, along with the effects on macrophages and platelets mentioned above.
In addition to its cardiovascular toxicity, TMAO Induces inflammation, oxidative stress, and fibrosis in renal parenchymal cells. Chronic administration of TMAO in animals has been shown to cause chronic tubulointerstitial fibrosis and progressive decline in renal function. (Tang et al. Circ Res, 2015)
In stark contrast to IS and PCS, TMAO Is hydrophilic and exhibits essentially zero protein binding. It is freely filtered by glomeruli and requires no active secretion by proximal tubular cells.
As described above, urea itself is a trivial uremic toxin at most, and through evolution, has become the solute of choice for nitrogen excretion by the kidneys after its formation in the liver and transit through the systemic circulation. While in isolation, elevated serum urea is relatively non-toxic, it certainly can contribute to gut dysbiosis in CKD.
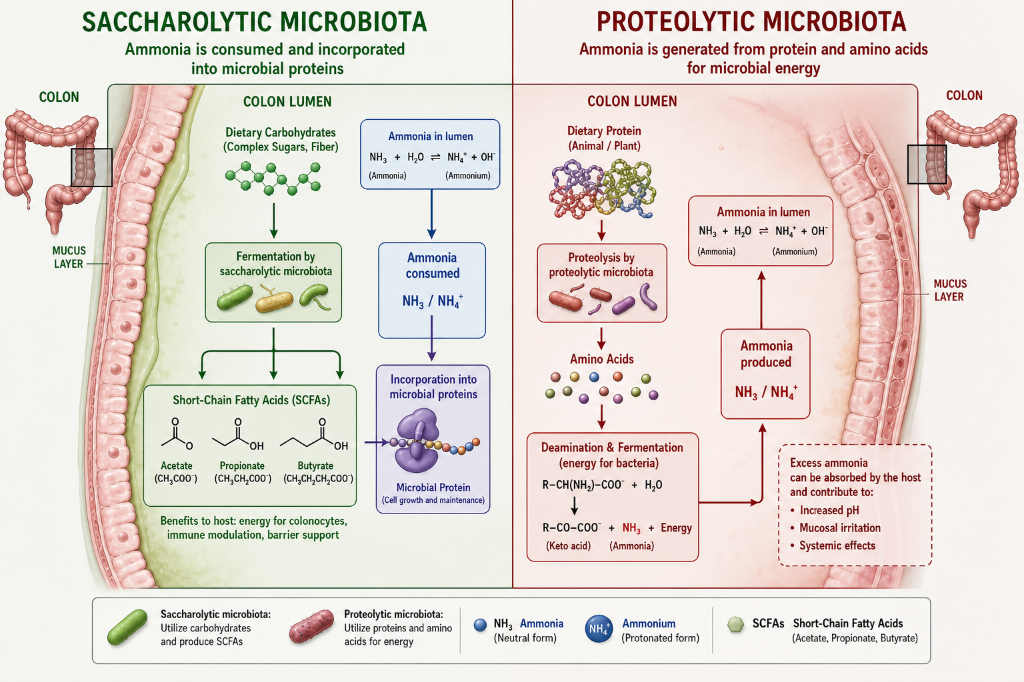
In CKD, as urea levels rise, urea diffuses across the gut lumen where it can be hydrolyzed back Into carbon dioxide and ammonia by bacterial urease enzymes. This ammonia generated within the gut lumen has a number of negative effects. It degrades the gut mucus layer which protects epithelial cells. It weakens the tight junctions between these epithelial cells which allows translocation of inflammatory mediators such as lipopolysaccharides to enter the circulation, resulting in a pro-inflammatory effect. The ammonia also raises intraluminal pH. This favors the growth of proteolytic, urease-producing bacteria, which produce more uremic toxins and more ammonia.
Keep in mind that the microbiome plays a key role in determining the fate of nitrogen (ammonia and others) present in the gut lumen. Saccharolytic microbes, fueled by carbohydrate from fiber, dispose of gut nitrogen by incorporating it into their own proteins and nucleic acids, which ultimately exit the body in feces; they serve as net nitrogen consumers. Conversely, proteolytic microbes, fueled by protein, deaminate proteins to use their carbon backbone for energy and therefore are net nitrogen producers.
Additionally, these proteolytic microbes produce urease which yields ammonia. After raising the pH in the gut lumen and thereby protecting the same proteolytic microbes that produced it, this same ammonia diffuses across the gut lumen, enters the circulation, and returns to the liver where it is then converted back into urea via the urea cycle.
Just as there are toxic, inflammatory gut microbial metabolites, Gut microbes are capable of producing beneficial, anti-inflammatory metabolites as well. Three groups of these will be discussed below. First are the short chained fatty acids (SCFAs) – acetate, propionate, and butyrate. These are produced by saccharolytic gut microbes via the fermentation of dietary fiber, in a ratio of roughly 1:1:3. They exert anti-inflammatory effects by promoting regulatory T-cell (Treg) expansion, increasing IL-10 levels, and reducing neutrophil recruitment. SCFAs, particularly butyrate, inhibit histone deacetylases (HDACs); this leads to a multitude of additional downstream anti-inflammatory effects. In addition, SCFAs are a preferred food source for colonocytes, generally helping maintain a strong colonic barrier function, and fostering a healthy, saccharolytic microbial milieu by maintaining an anaerobic, low pH colonic environment (see below).
Next are secondary bile acids. These are metabolic products of gut microbes Acting on primary bile acids which are synthesized in the liver and enter the gut lumen in bile. The steps involved in the formation of secondary bile acids and the mechanisms by which they anti-inflammatory effects follows.
Two primary bile acids are synthesized in the liver from cholesterol – cholic acid (CA) and chenodeoxycholic acid (CDCA). To make them more soluble in water, these primary bile acids are conjugated with an amino acid, either glycine or taurine, yielding the 4 conjugated primary bile acids that are stored in the gallbladder and secreted into the duodenum after a meal – glycocholic acid, taurocholic acid, glycochenodeoxycholic acid, and taurochenodeoxycholic acid. About 95% of bile acids are resorbed in the terminal ilium And return to the liver via the portal circulation, where they are resecreted as bile (enterohepatic circulation). Five percent of primary bile acids escape resorption in the ilium and enter the colon. This is the substrate pool that is acted on by the microbiome.
In order to form anti-inflammatory secondary bile acids, primary bile acids must undergo two steps that are mediated by gut microbes – bile acid deconjugation, which is the gateway step, and 7α-dehydroxylation, which is the key step and produces anti-inflammatory secondary bile acids. Primary bile acid deconjugation is considered the gateway step In secondary bile acid formation because a diverse group of gut microbes have this capability. In contrast, the second or key step, 7α-dehydroxylation, is a multistep process carried out by a relatively small group of microbes, Clostridium Scindens being the first discovered and most studied. Microbial strains that are capable of 7α-dehydroxylation only thrive in a saccharolytic strictly anaerobic ecosystem, where SCFAs help to maintain a low pH.
Once formed within a colonic lumen, secondary bile acids themselves suppress the growth of proteolytic gram negative enteric bacterial species via cell membrane disruption. Secondary bile acids are the most hydrophobic form of bile acids, which gives them the greatest capacity to disrupt cellular membranes. Unlike the apical membranes of colonocytes and membranes of bacterial species capable of 7α-dehydroxylation, gram negative enteric bacteria have no receptors or transporters for secondary bile acids, which makes them particularly sensitive to cell membrane disruption by secondary bile acids.
After being formed in the colonic lumen, secondary bile acids, binding to two distinct receptor types – TGR5 and FXR, exert downstream anti-inflammatory effects in multiple tissues and organs.
Secondary bile acids pass through the colonic epithelium, enter the portal circulation, and enter hepatocytes. Here they are reconjugated, secreted in bile, and bind to epithelial cells in the ileum.
A small quantity avoids first pass metabolism in the liver and enters the systemic circulation, and reaches the kidney, exerting downstream anti-inflammatory effects In all these tissues along the way.
There is a “therapeutic window of benefit” with regard to secondary bile acid levels. Proteolytic gram negative bacteria are particularly prone to membrane disruption at moderate levels. But as levels continue to increase, particularly of DCA levels, host cellular structures and organelles begin to experience membrane disruption and damage as well, causing a DAMP-type inflammatory cascade. In the presence of an extremely high fat diet, the microbiome produces high levels of DCA, causing this DAMP-type inflammatory response in colonic epithelial cells. When biliary obstruction or cholestasis occurs, DCA accumulates In hepatocytes, causing the same DAMP response in them.
Third are protective indole metabolites. As discussed above, dietary tryptophan not absorbed in the small intestine can be metabolized to indole by proteolytic gut enterobacteriaceae, which leads to downstream formation of indoxyl sulfate, a powerful uremic toxin. This is obviously a pathogenic pathway. However, when healthy, saccharolytic gut microbiotal conditions exist, this same dietary tryptophan can be metabolized by two other distinct pathways by gut microbes which lead to beneficial metabolites.
One beneficial pathway (IPA pathway), first described in Clostridium Sporogenes and since described in several other microbes, involves the metabolism of tryptophan to indole-3-propionic acid (IPA) and indole-3-lactic acid (ILA). (Dodd et al, Nature, 2017) The second beneficial pathway Involves certain Lactobacillus species, which metabolize tryptophan into indole-3-aldehyde (IAld).
As with other beneficial gut microbiome metabolites, these three molecules (IPA,ILA, and IAld) exert downstream anti-inflammatory effects. Of the three, IPA Is the most potent and significant, escaping hepatic first-pass metabolism And exerting significant systemic effects. IPA levels serve as a biomarker of gut microbiome health, with IPA levels correlating inversely with IS and PCS levels, as well as with cardiovascular risk and CKD progression.
Dietary fiber
The theme of benefit from a fiber-rich diet and healthy saccharolytic microbiome runs through virtually every example of gut-derived toxic and beneficial metabolites mentioned above, so a more complete description of this fiber and microbial ecosystem is warranted. This ecosystem can be conceptualized as a forest filled with a variety of plant life (fiber types) and a variety of different animals (microbe types).
Fiber comes in many forms, with each form having a subset of microbiomes capable of metabolizing it. Some fiber types include the following:
- Resistant starches are glucose polymers in crystalline granule form.
- Cellulose is a β-1,4-linked glucose polymer. This is the most abundant organic molecule on earth. It is largely non-fermentable in humans due to limited colonic cellulolytic capacity.
- Hemicelluloses are a major component of plant cell walls. This group includes arabinoxylans – a major component of wheat, rye, and rice brans. The group also includes galactomannans such as guar gum and locust bean gum, which are fermentable thickeners and emulsifiers.
- Pectins are galacturonic acid-rich polymers with complex branching, abundant in fruits and vegetables.
- Inulin and Fructooligosaccharides (FOS) are β-2,1-linked fructose polymers of varying chain length. Inulin is long (10-60); FOS short (2-10) These are found in chicory, onion, garlic, asparagus, artichoke.
- Galactooligosaccharides (GOS) are galactose polymers found in legumes. They are also produced from lactose commercially.
- Beta-glucans are glucose polymers with mixed β-1,3 and β-1,4 linkages. These are found in oats and barley.
- Miscellaneous fibers (resistant dextrins, psyllium, and others) each have distinct structural features requiring specific enzymatic attack.
Degrading these diverse fiber forms requires a variety of microbes, working in concert. As fiber is degraded, the products of one phase become the substrates of a subsequent phase. A critical end-product of gut microbial metabolism is butyrate, which is the preferred energy source for colonic epithelial cells. The hierarchical structure of the saccharolytic microbiome and the concept of cross feeding are discussed next.
Level 1 – the complex polymer degraders
These organisms convert large, complex polymers into oligosaccharides. The prototype here is Bacteroides thetaiotaomicron and related Bacteroides species. The genome of B. thetaiotaomicron contains more than 80 dedicated gene complexes, called polysaccharide utilization loci (PULs), for digesting specific types of complex fiber polymers. This microbe attacks pectins, xylans, mannans, various hemicelluloses, and mucin glycans (it can also degrade host mucus when dietary fiber is absent). These Bacteroides species perform initial hydrolysis of large polymers at their cell surface and subsequently import a portion of the oligosaccharide fragments into the cell for further metabolism. However, many of these oligosaccharide fragments remain outside the cell in the surrounding environment and become substrates for the next layer of microbes.
Ruminococcus bromii Is another important microbe on this level. This is a clostridial species with the unique ability to degrade resistant starch granules. Because the ability to breakdown resistant starch completely collapses in the absence of R. bromii, It is considered a keystone microbial species. As with the case of the Bacteroides species mentioned above, R. bromii produces amylases on its cell surface that attach to and begin breaking down resistant starch granules. Again, some of the oligosaccharides and monosaccharides produced are taken up by the Ruminococci themselves, while the rest remain in the local environment to serve as substrates for the next level.
Level 2 – the oligosaccharide specialists
These organisms cannot attack intact polymers directly but are highly efficient at capturing and fermenting the oligosaccharides and simple sugars released by Level 1. Bifidobacteria species are the prototype here. Bifidobacteria ferment sugars through a unique metabolic pathway that splits hexose phosphate asymmetrically to produce acetate and lactate. Again, these fermentation products function as substrates for the next microbe layer.
Eubacterium rectale ferments maltose, maltotriose, maltooligosaccharides, and short-chain glucose oligomers to butyrate. This makes it somewhat unique, as butyrate is generally a product of level 3.
Roseburia species also bridge levels 2 and 3 by fermenting oligosaccharides into butyrate. Roseburia also has some level 1 activity, as it is able to break down some large, complex polymer types.
Level 3 – the butyrate producers
These organisms convert acetate and lactate to butyrate. The prototype here is Faecalibacterium prausnitzii, which is the single most abundant bacterium in the healthy human colon. F. prausnitzii is an acetate-consuming, sugar-fermenting butyrate producer, deriving its energy from the fermentation of simple sugars and some oligosaccharides, and forming butyrate as an electron sink, which regenerates NAD+ and other oxidized intermediates. The regeneration of these oxidized intermediates allows fermentation to continue. F. prausnitzii also possesses an extracellular electron shuttle system which allows it to tolerate (and even benefit from) the trace oxygen present at the mucosal surface. Being positioned in this microaerobic zone of the mucus layer, close to the epithelium, facilitates delivery of butyrate to colonic epithelial cells.
Anaerostipes caccae and related species specialize in converting lactate-to-butyrate. Solely through this conversion, the organisms are able to extract adequate energy and sustain growth. In addition to providing butyrate for colonic epithelial cells, this process prevents the accumulation of excess lactate, which could lower pH too far, cause mucosal irritation, and in extreme cases contribute to D-lactic acidosis.
Level 4 – the hydrogen gas consumers
Hydrogen gas is an end product of fermentation in levels 1-3. Excessive accumulation of hydrogen gas (H₂) can make NADH-oxidizing reactions thermodynamically unfavorable, thereby halting any further fermentation. Organisms that consume H₂ are essential for keeping the H₂ partial pressure low enough for fermentation to continue. Gut microbes use hydrogen gas to form three different reduction products – acetate, hydrogen sulfide, and methane. Each of these products has different effects on the metabolism and health of the colon.
The beneficial end product of the three is acetate. The prototype acetogenic microbial group is Blautia hydrogenotrophica and related species. Acetogens convert hydrogen gas and carbon dioxide into acetate plus water. This acetate can then be metabolized by the butyrate producers of level 3 to form butyrate, which feeds the colonic epithelium.
The modestly detrimental end product is methane. The prototype methanogenic microbial group is Methanobrevibacter smithii and related archaea species. Methanogens convert hydrogen gas and carbon dioxide to form methane and water. Methane is either expelled as flatus or absorbed into the bloodstream and then exhaled via the lungs. The detrimental effect of methane is that it inhibits smooth muscle contractility which results in increased intestinal transit time (discussed below). This creates a vicious cycle of slower transit – more fermentation time – more methanogenesis – even slower transit time – and so on.
Hydrogen sulfide is the most potentially detrimental or toxic end product. The prototype sulfate-reducing microbes are the Desulfovibrio species, which use hydrogen gas to reduce sulfate anions forming hydrogen sulfide and water. Hydrogen sulfide (H₂S), in high concentrations, is toxic to colonocytes by blocking mitochondrial metabolism, including butyrate oxidation. The sulfur-containing amino acids, cysteine and methionine, are present in significantly higher concentrations In animal proteins than in plant proteins. These amino acids are a highly bioavailable source of sulfate for sulfate-reducing microbes. Therefore, a high animal protein diet correlates with higher hydrogen sulfide production.
Interestingly, at low concentrations, hydrogen sulfide is nontoxic to colonocytes. They possess the biological machinery to oxidize hydrogen sulfide and derive energy., in the presence of high levels, this machinery is overwhelmed and the H₂S becomes a mitochondrial toxin.
Level 5 – the metabolic specialists
These are organisms that perform specific, high-value metabolic functions (secondary bile acid production, IPA production, and IAld production), which we have discussed above. They do not degrade fiber themselves. but depend on the saccharolytic ecosystem provided by levels 1-4.
To review, Clostridium scindens and related species produce beneficial secondary bile acids. These organisms benefit from a strictly anaerobic environment and low colonic pH, Butyrate, produced by saccharolytic microbiome metabolism, is oxidized by colonocytes, which helps to maintain a strictly anaerobic environment. Also, along with other SCFAs, butyrate helps maintain a low PH in the colonic lumen.
Clostridium sporogenes, which produces the beneficial indole metabolite indole-3-propionic acid (IPA). This is actually a proteolytic microbe, but it is an obligate anaerobe. The low pH and anaerobic environment provided by a saccharolytic microbial ecosystem provides a competitive advantage over other proteolytic microbes.
Lactobacillus reuteri and related species produce indole-3-aldehyde, another beneficial indole metabolite. These microbes benefit from sugars and oligosaccharides produced in level 1, as well as the acidic pH produced by the saccharolytic microbe community.
Level 6 – the controlled minority – enterobacteriaceae
These are gram negative facultative anaerobes, with the prototype enterobacteriaceae being Escherichia coli (E. coli). In a healthy saccharolytic microbial environment, enterobacteriaceae comprise less than 1 percent of the gut microbial population. However, in a dysbiotic milieu, the population of enterobacteriaceae can expand significantly, reaching up to 15% chronic disease states end up to 90% in severe illness. As part of their normal life cycle, all enterobacteriaceae shed lipopolysaccharides (LPS) from their outer membrane. Lipopolysaccharides are highly immunostimulatory and pro-inflammatory, thus making all enterobacteriaceae, to some degree, intrinsically pro-inflammatory.
While enterobacteriaceae are facultative anaerobes, aerobic conditions are much more favorable for their growth. Also, while they can tolerate a relatively low pH of a saccharolytic environment, a higher pH, as is found in a proteolytic gut milieu, favors their growth. Enterobacteriaceae are quite metabolically flexible. As described extensively above, enterobacteriaceae are capable of using proteins and amino acids for energy, which leaves ammonia behind is a waste product. However, their preferred energy sources are simple sugars and short oligosaccharides, as is found in a healthy, saccharolytic gut microbiome. In this scenario, nitrogen (ammonia) Is incorporated from the gut lumen to form amino acids and ultimately bacterial proteins and is ultimately excreted as microbial biomass in feces.
To review, a saccharolytic microbiome controls the enterobacteriaceae population through a number of mechanisms that work synergistically. First, their energy source of choice, sugars and short oligosaccharides, are available but in somewhat limited supply because of competition with other saccharolytic microbes. Second, SCFAs produced from saccharolytic metabolism are weak acids and lower the pH in the colonic lumen. SCFAs also enter microbial cells, which lowers the microbial intracellular pH. Since enterobacteriaceae prefer a normal or alkaline pH, this acidification serves as a metabolic burden and limits their growth. Additionally, SCFAs are oxidized by colonocytes. This reduces oxygen tension in the colonic lumen and prevents aerobic metabolism, the preferred mode of energy production for Enterobacteriaceae.
Third, secondary bile acids have a direct membrane disrupting effect on enterobacteriaceae. Secondary bile acids (SBAs) are produced by the keystone clostridial species that perform 7α-dehydroxylation of primary bile acids. These microbes can only grow in a healthy saccharolytic gut environment. Unlike many microbes in the saccharolytic microbiome, enterobacteriaceae have no specific metabolic defenses against secondary bile acids, and therefore their numbers are selectively depressed In the presence of SBAs. Lastly, some saccharolytic microbial species produce antimicrobial chemicals and small peptides which place further selective pressure on enterobacteriaceae. In addition to these microbe-produced antimicrobial chemicals, colonocytes produce antimicrobial molecules which are upregulated by SCFAs and SBAs, which places additional selective pressure on enterobacteriaceae.
Transit time through the colon plays a significant role in gut microbiome composition. In humans, whole gut transit time usually varies between 24 and 72 hours, with most of the variability occurring in the colon. As material traverses through the colon fermentable fiber is progressively consumed and water and electrolytes are resorbed. Once fermentable fiber is depleted, proteolytic microbes thrive, pH rises SCFA production goes down and ammonia production goes up.
If transit time is excessively slow, fermentable fiber depleted at a more proximal point in the colon. This allows for more time and colonic surface area for proteolytic metabolism to take over, resulting in increased production of toxic and pro-inflammatory metabolites as well as a weakening of the colonic epithelial barrier.
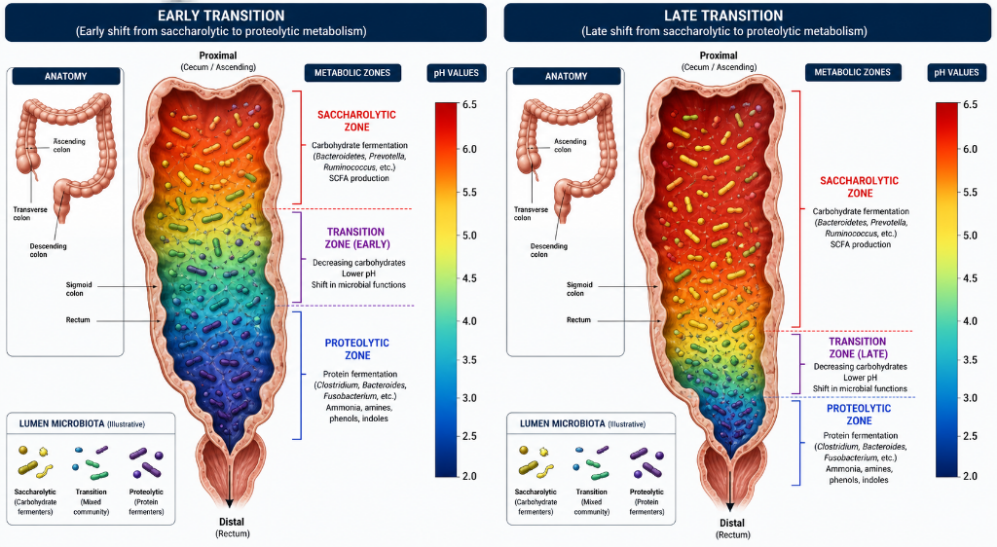
Conversely, if transit time is too rapid, not all fermentable fiber will be metabolized into beneficial, anti-inflammatory metabolites, resulting in a loss of some metabolic benefit. So, an ideal transit time would have depletion of fermentable fiber occurring at a more distal colonic segment. The association between a relatively short transit time and a saccharolytic microbiome has been shown in humans, regardless of diet or dietary fiber. (Roager et al. Nature Microbiology, 2016)
References
Eknoyan G et al. (1969) Platelet function in renal failure NEJM 280(13) p.677-81
Johnson WJ et al. (1972) Effects of urea loading in patients with far-advanced renal failure Mayo Clinic Proc_47(1) p.21-9
Aronov PA et al. (2011) Colonic contribution to uremic solutes JASN 22(9) p.1769-76
Einheber A & Carter D (1966) Microbial flora in uremia, survival times of germ-free and conventionalized rats after bilateral nephrectomy J Exp Med, 123(2) p.239-50
Tang WW et al. (2015) Gut microbiota-dependent TMAO pathway contributes to both development of renal insufficiency and mortality risk in CKD Circ Res 116(3) p.448-55
Dodd D et al. (2017) A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites Nature 551(7682) p.648-52
Roager, HM et al. (2016) Colonic transit time is related to bacterial metabolism and mucosal turnover in the gut Nature microbiology 1(9) 16093